Super-Resolution Imaging of ESCRT-Proteins at HIV-1 Assembly Sites
24-Feb-2015
PLOS Pathogens, 2015, DOI: 10.1371/journal.ppat.1004677, published on 24.02.2015
PLOS Pathogens, online article
PLOS Pathogens, online article
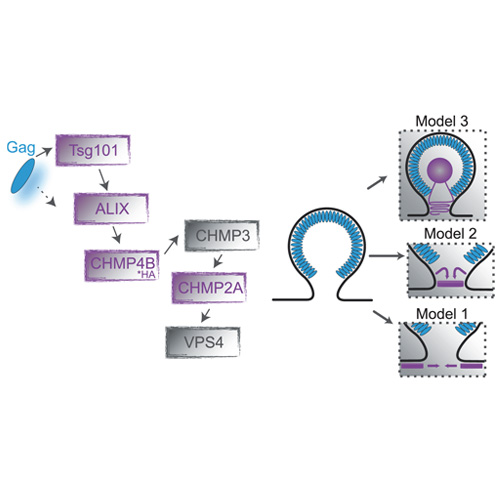
The cellular endosomal sorting complex required for transport (ESCRT) machinery is involved in membrane budding processes, such as multivesicular biogenesis and cytokinesis. In HIV-infected cells, HIV-1 hijacks the ESCRT machinery to drive HIV release. Early in the HIV-1 assembly process, the ESCRT-I protein Tsg101 and the ESCRT-related protein ALIX are recruited to the assembly site. Further downstream, components such as the ESCRT-III proteins CHMP4 and CHMP2 form transient membrane associated lattices, which are involved in virus-host membrane fission. Although various geometries of ESCRT-III assemblies could be observed, the actual membrane constriction and fission mechanism is not fully understood. Fission might be driven from inside the HIV-1 budding neck by narrowing the membranes from the outside by larger lattices surrounding the neck, or from within the bud. Here, we use super-resolution fluorescence microscopy to elucidate the size and structure of the ESCRT components Tsg101, ALIX, CHMP4B and CHMP2A during HIV-1 budding below the diffraction limit. To avoid the deleterious effects of using fusion proteins attached to ESCRT components, we performed measurements on the endogenous protein or, in the case of CHMP4B, constructs modified with the small HA tag. Due to the transient nature of the ESCRT interactions, the fraction of HIV-1 assembly sites with colocalizing ESCRT complexes was low (1.5%-3.4%). All colocalizing ESCRT clusters exhibited closed, circular structures with an average size (full-width at half-maximum) between 45 and 60 nm or a diameter (determined using a Ripley’s L-function analysis) of roughly 60 to 100 nm. The size distributions for colocalizing clusters were narrower than for non-colocalizing clusters, and significantly smaller than the HIV-1 bud. Hence, our results support a membrane scission process driven by ESCRT protein assemblies inside a confined structure, such as the bud neck, rather than by large lattices around the neck or in the bud lumen. In the case of ALIX, a cloud of individual molecules surrounding the central clusters was often observed, which we attribute to ALIX molecules incorporated into the nascent HIV-1 Gag shell. Experiments performed using YFP-tagged Tsg101 led to an over 10-fold increase in ESCRT structures colocalizing with HIV-1 budding sites indicating an influence of the fusion protein tag on the function of the ESCRT protein.